
DATE2026.05.11 #Press Releases
転写抑制を調節する新規因子の発見
ータンパク質の安定性制御を介した転写抑制のしくみー
発表のポイント
- 植物を用いた研究から、真核生物に広く保存されたヒストン修飾H3K9ジメチル化による転写抑制に必要な新規因子MBD8を同定し、その機能を解明しました。
- MBD8は転写抑制に働くLDL2のタンパク質安定性を高めることで、転写抑制を促進することがわかりました。
- MBD8は多くの植物に存在するため、植物の高い環境適応能力の基盤となる遺伝子発現制御機構解明の一助となることが期待されます。
1.jpg)
MBD8はLDL2のタンパク質安定性を高め転写抑制に働く
概要
東京大学大学院理学系研究科の森秀世特任研究員 (研究当時)、稲垣宗一准教授 (研究当時)、角谷徹仁東京大学名誉教授 (研究当時:教授) らによる研究グループは、植物のシロイヌナズナ (注1) を用いた研究から、新規因子MBD8がH3K9ジメチル化による転写抑制に必要であることを明らかにしました。MBD8は転写抑制に働くLDL2のタンパク質安定性を高めることで、転写抑制を促進します。このMBD8の機能は他の植物に保存されていることが予想され、本研究の成果は、植物が転写を抑制する分子機構とそれにより制御される発生過程や環境応答メカニズムへの理解に役立つことが期待されます。
発表内容
ヒストンH3タンパク質の9番目のリジンのジメチル化 (H3K9me2) は、真核生物に広く保存されたヒストン修飾(注2)であり、不必要な遺伝子の転写を抑制することが知られています。しかしながら、H3K9me2がどのようにして転写を抑制しているのか、そのメカニズムには未解明な点が多く残されていました。
この疑問に答えるため、私たちは植物のシロイヌナズナを用いて研究を行ってきました。その結果、H3K9me2はヒストン脱メチル化酵素 (注3) LDL2を呼び込み、別のヒストン修飾であるH3の4番目のリジンのモノメチル化 (H3K4me1) を除くことによって転写を抑制していることが明らかになりました (関連プレスリリース①)。H3K4me1には転写を促進する働きがあるため (関連プレスリリース②)、H3K4me1を除くことで転写を抑制できるということです。本研究ではLDL2の機能に必要な新規因子としてMBD8を同定し、その機能を明らかにしました。
本研究ではまず、LDL2に結合して共に働く因子を探索するため、免疫沈降質量分析 (IP-MS) (注4) を行いました。その結果、MBD8というタンパク質がLDL2と結合することがわかりました。MBD8はメチル化シトシン (注5) に結合するMethyl-CpG Binding Domain (MBD) を持つタンパク質ファミリーに属しますが、その機能は明らかになっていませんでした。本研究チームは、MBD8がLDL2の機能に必要なのではないかと仮説をたて検証しました。
ibm1変異体では、本来存在すべきではない遺伝子領域にH3K9me2が広がり、LDL2によるH3K4me1除去を引き起こされることで、不適切な遺伝子抑制とそれに伴う発生異常が生じます。一方ibm1 ldl2二重変異体では、LDL2によるH3K4me1除去が起こらないため、発生異常が抑えられます。もしMBD8がLDL2の機能に必要ならば、ibm1 mbd8二重変異体でも同様に発生異常が抑えられるのではないかと考えました。実際、ibm1 mbd8二重変異体では発生異常が抑えられ (図1)、ibm1変異体に比べてH3K4me1レベルと転写量も回復していました。このことから、MBD8はLDL2がH3K4me1を除く機能に必要な因子であることがわかりました。
2.jpg)
図1:MBD8はH3K9me2による転写抑制に必要である
ibm1変異体では過剰な遺伝子抑制により発生異常が見られるが、ibm1 mbd8二重変異体ではその発生異常が抑えられる。
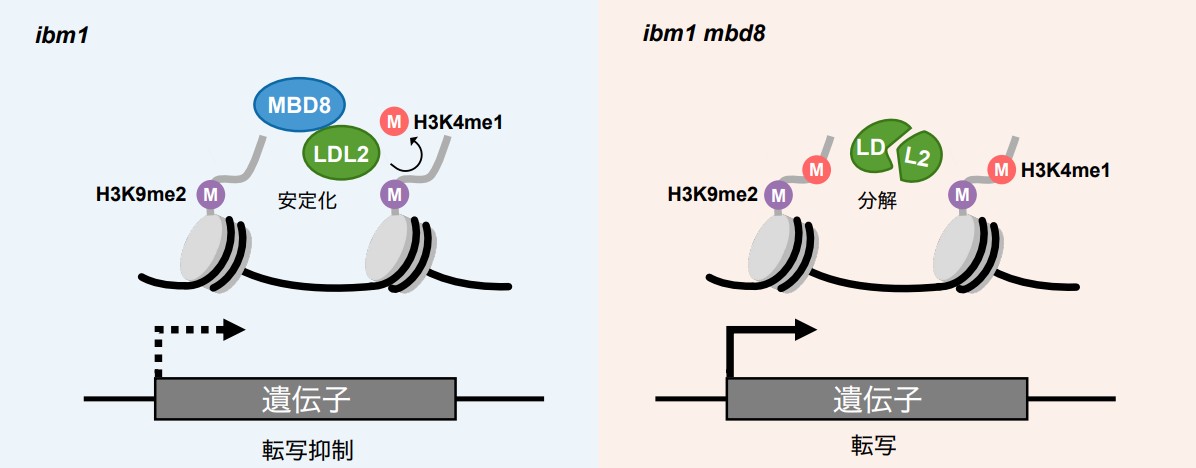
さらにMBD8の機能を調べた結果、MBD8はLDL2のタンパク質安定性を保つのに重要であることがわかりました。mbd8変異体では、LDL2タンパク質が分解されてしまい、その結果LDL2による転写抑制が正常に行われなくなります (図2)。興味深いことに、MBD8のMBDはメチル化シトシン結合能を失っており、MBDを欠いたMBD8タンパク質でもLDL2による転写抑制を引き起こす能力があることがわかりました。これらの結果は、MBD8が従来想定されていた機能とは異なる、独自の役割を持つことを示しています。シロイヌナズナ以外の植物においても、MBD8とLDL2の結合に重要なアミノ酸領域は保存されています。本研究で明らかになった転写抑制機構は、植物に共通する仕組みである可能性が高く、植物の柔軟な発生制御や高い環境適応能力の理解に貢献することが期待されます。
3.jpg)
図2:本研究で明らかにしたMBD8とLDL2による転写抑制モデル
ibm1変異体ではH3K9me2が蓄積した遺伝子においてLDL2がH3K4me1を除き、転写が抑制される。MBD8はLDL2のタンパク質安定化に働く。ibm1 mbd8二重変異体ではLDL2が安定化されず分解されるため、H3K4me1が除かれず転写抑制されない。
〇関連情報:
「プレスリリース①遺伝子転写の促進と抑制の「二刀流」」(2024/06/27)
「プレスリリース②転写活性を植物ゲノムに記録する機構」(2023/10/20)
発表者・研究者等情報
東京大学
大学院理学系研究科 生物科学専攻
森 秀世 特任研究員(研究当時)
稲垣 宗一 准教授(研究当時)(現:京都大学大学院理学研究科 教授)
角谷 徹仁 教授(研究当時)(現:東京大学名誉教授)
論文情報
-
雑誌名 Nucleic Acids Research 論文タイトル MBD8 is required for LDL2-mediated transcriptional repression downstream of H3K9me2 in Arabidopsis著者 Shusei Mori*, Akihisa Osakabe, Juliarni, Yuriko Tanaka, Mami Hirayama, Soichi Inagaki*, Tetsuji Kakutani* (*責任著者) DOI番号 10.1093/nar/gkag361
研究助成
本研究は、科研費(課題番号:21H04977、23H00365、24K21268、20H05913、23K23565、24K21926、25K02252、24H0135、24K02077、24K02002、25K02258、25H02544、25H01300)、HFSP(RGP0025/2021)の支援により実施されました。
用語解説
注1 シロイヌナズナ
代表的な実験植物。シロイヌナズナはゲノムサイズが小さく、世代時間が短く、遺伝子組換え等も容易で、また、真核生物に存在するヒストン修飾のほとんどを持つため、ヒストン修飾制御の研究に適している。↑
注2 ヒストン修飾
ヒストンはゲノム中でDNAが巻き付いているタンパク質複合体で、その翻訳後修飾(メチル化、アセチル化、リン酸化など)は局所的なDNAの高次構造などに変化をもたらす。エピジェネティックな遺伝子制御に関わる修飾である。↑
注3 ヒストン脱メチル化酵素
ヒストンの特定のアミノ酸からメチル化を除去する酵素を脱メチル化酵素と呼ぶ。代表的な脱メチル化酵素にLSD1とJUMONJI型タンパク質がある。LDL2はLSD1型タンパク質、IBM1はJUMONJI型タンパク質である。↑
注4 免疫沈降質量分析(IP-MS)
抗体によって標的タンパク質とその結合タンパク質をプルダウンする。続く質量分析でアミノ酸一次配列を決定することで、標的タンパク質と結合するタンパク質を網羅的に探索できる。↑
注5 メチル化シトシン
シトシンはDNAを構成する塩基のうちの一つ。このシトシンにメチル基が付加されたものをメチル化シトシンと呼び、転写抑制に働くことが知られている。↑