
DATE2022.02.14 #Press Releases
植物ゲノムに不活性領域を作り出す機構
藤 泰子(生物科学専攻 助教)
角谷 徹仁(生物科学専攻 教授)
発表のポイント
- ゲノム(注1)を不安定化する増殖性の配列は、DNAをメチル化(注2) することで沈静化されている。植物における2種類のDNAメチル化経路が局所的に増強しあうことを明らかにした。
- 一方、ゲノム全体では、この機構が暴走しないように抑える仕組みがあることを見出した。
- ゲノムの局所的および全体で逆方向に働く仕組みにより、強力に不活性化された領域を作り、ゲノムの不安定化を防いでいると考えられる。
発表概要
動物や植物のゲノムは可動性で増殖する性質のDNAを多量に含み、これがゲノムの不安定化や癌などの疾病の原因になる。増殖性配列を不活性化する機構として植物や脊椎動物ではDNAのメチル化が知られている。しかしながら、ゲノム中の特定のDNA配列だけをメチル化し不活性化する仕組みについては未解明の点が多い。
東京大学大学院理学系研究科の藤泰子助教と角谷徹仁教授らのグループは、シロイヌナズナという植物の変異体を用いた研究の結果、DNAメチル化の間の局所的な正のフィードバックと、ゲノム全体での負のフィードバックを見出した。この両方の経路により、働くべき遺伝子を働かせながら、不活化領域に限定した抑制が可能となり、ゲノムの不安定化を防いでいると考えられる。
発表内容
増殖性の配列である転移因子(transposable element: TE)は、脊椎動物や陸上植物のゲノムの大部分をしめ、ゲノム進化に大きく貢献する。その一方で、TEはその潜在的な転移能力からゲノム安定性の脅威となる。このため、染色体の主要な構成タンパク質であるヒストンタンパク質の特異的な翻訳後修飾(注3) やDNAのメチル化などの複数のエピジェネティックな機構(DNA配列以外の形で遺伝子のON/OFF情報を決め、細胞分裂後にも継承される機構)により抑制されている。
植物のゲノムにおいては、DNA メチル化による抑制が重要である。一般にシトシンのメチル化は、グアニン(G)の直前にあるシトシン(CpG)に多く、このようなCpG配列のメチル化(mCG)は、維持メチル化という機構によって細胞分裂後にまでメチル化の有無が継承される(注4) 。植物で見られるCpG以外の配列のシトシンメチル化(mCH; HはG以外の塩基)は、これを触媒する酵素がヒストンH3タンパク質の9番目のリジン残基のメチル化(H3K9me)を多く含む領域にリクルートされ、また、H3K9meを触媒する酵素がmCHを多く含む領域にリクルートされる相互依存的関係を持つため、これらの修飾も染色体の分裂後に継承され、増強される。一方でmCHとmCGは、ほぼ独立に継承されており、この両者が共にTEを多く含む不活性領域に集中する仕組みはわかっていなかった。
本研究では、mCHとH3K9meを特定のゲノム領域に新たに確立する際に、mCGが大きく貢献することを見出した(注5) 。意外なことに、mCGがゲノム全体で低下した条件では、この経路が強力に働く。その他の幾つかの変異体を用いた結果も、ゲノム全体のDNAメチル化が、mCH誘発を負に制御するというフィードバックの存在を支持した(図1)。
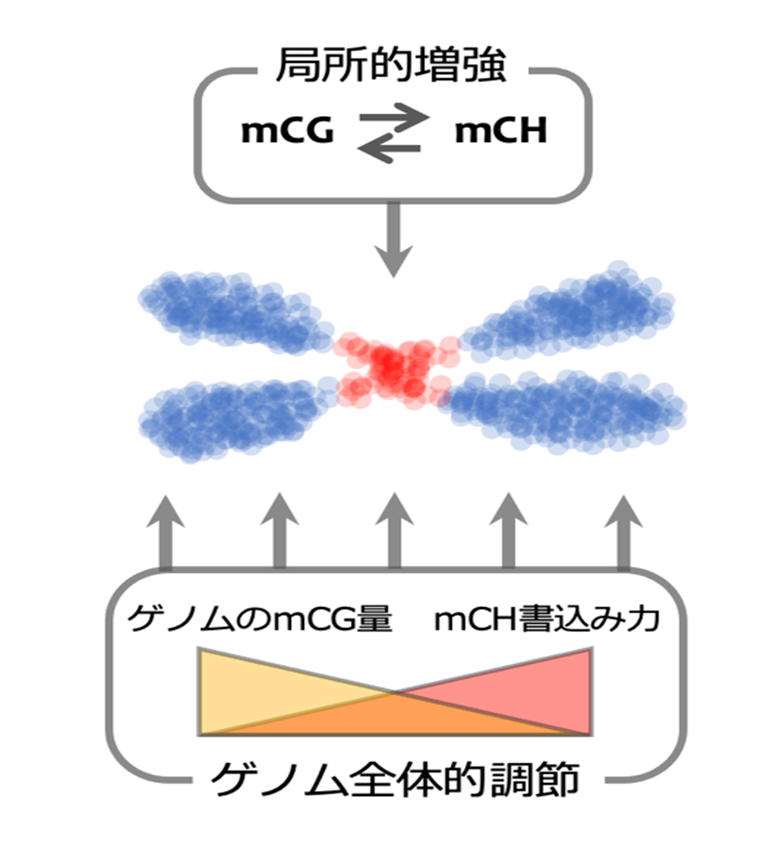
図1:植物ゲノムに不活性化領域を作り出す機構
植物ゲノムはトランスポゾンを多く含む不活性領域(赤)と遺伝子を多く含む活性領域(青)に大別され、それぞれ固有のエピジェネティック修飾を持つ。本研究により、2種のDNAメチル化(mCGおよびmCH)の間で局所的な正の相互作用およびゲノム全体での負の相互作用が見出された。この二つの相互作用により、ゲノムにおいて適切に不活性化領域が形成されると考えられる。
DNAメチル化やH3K9meによる抑制を仲介する因子の多くが植物のみならず動物にも保存されているため、本研究により明らかとなった機構が、他の生物にも保存されている可能性がある。今後は、この新たな経路の分子機構解明が、大きな波及効果を持つと期待される。
発表雑誌
-
雑誌名 Nature Communications 論文タイトル Local and global crosstalk among heterochromatin marks drives DNA methylome patterning in Arabidopsis 著者 Taiko Kim To*, Chikae Yamasaki, Shoko Oda, Sayaka Tominaga, Akie Kobayashi, Yoshiaki Tarutani, Tetsuji Kakutani* DOI番号
用語解説
注1 ゲノム
特定の生物の持つ遺伝情報全体、あるいはその分子的実体である1セットのDNA のこと。↑
注2 DNAのメチル化
動物や植物ではDNA中のシトシンの5位がメチル化されることがある。このようなメチル化が転写開始点付近にあると、転写が抑制されていることが多い。Gの直前のシトシンのメチル化(mCG)に加え、植物ではG以外の塩基(H:A, T, あるいはC)の直前のメチル化(mCH)も重要である。 ↑
注3 翻訳後修飾
タンパク質の中には翻訳の後にメチル化やリン酸化などの修飾を受けるものがある。このような修飾がタンパク質の機能を制御することがある。 ↑
注4 DNAメチル化の「継承」
遺伝子のON/OFF状態が細胞分裂後に継承されることがエピジェネティックな現象の根幹であり、この分子基盤の一つがDNAメチル化の継承である。多くの動植物は、CG配列の片鎖にメチル化がある場合に、もう一方の鎖のCG配列をメチル化する酵素を持つ。この酵素は、維持メチル化酵素と呼ばれ、その活性により、DNAの複製時に新たに合成された鎖のCG配列もメチル化される。このような機構により、両鎖がメチル化されたCG配列はDNA複製後も両鎖のメチル化が「継承」される。一方で、植物におけるmCHはH3K9meを介した別の経路により継承される。 ↑
注5 DNAメチル化の「確立」
上記の「継承」に加え、メチル化のなかったシトシンが新たにメチル化される仕組みがあり、これをDNAメチル化の「確立」と呼ぶ。本研究では、DNAメチル化に必要な因子を欠損した変異体下でDNAメチル化を喪失させた後、この変異体に機能する遺伝子を導入することで、「確立」活性を検出した。 ↑