
DATE2025.05.12 #Press Releases
細菌のゲノム構造進化を実験室で観測
――トランスポゾンに駆動された進化を加速する新手法を開発――
発表のポイント
- 高活性なトランスポゾンを大腸菌ゲノムに導入することで構造変異の頻度を高め、ゲノム構造の進化を加速させることに成功しました。
- 従来、数十年かけて観測されてきたゲノム構造の進化と同等の進化をわずか10週間で再現した上、新たなトランスポゾンの出現する過程まで観測できました。
- 本技術のさらなる発展を通じて、共生細菌や病原性細菌が出現する進化過程の理解が進むほか、産業上重要な細菌の改良の高速化に貢献すると期待されます。
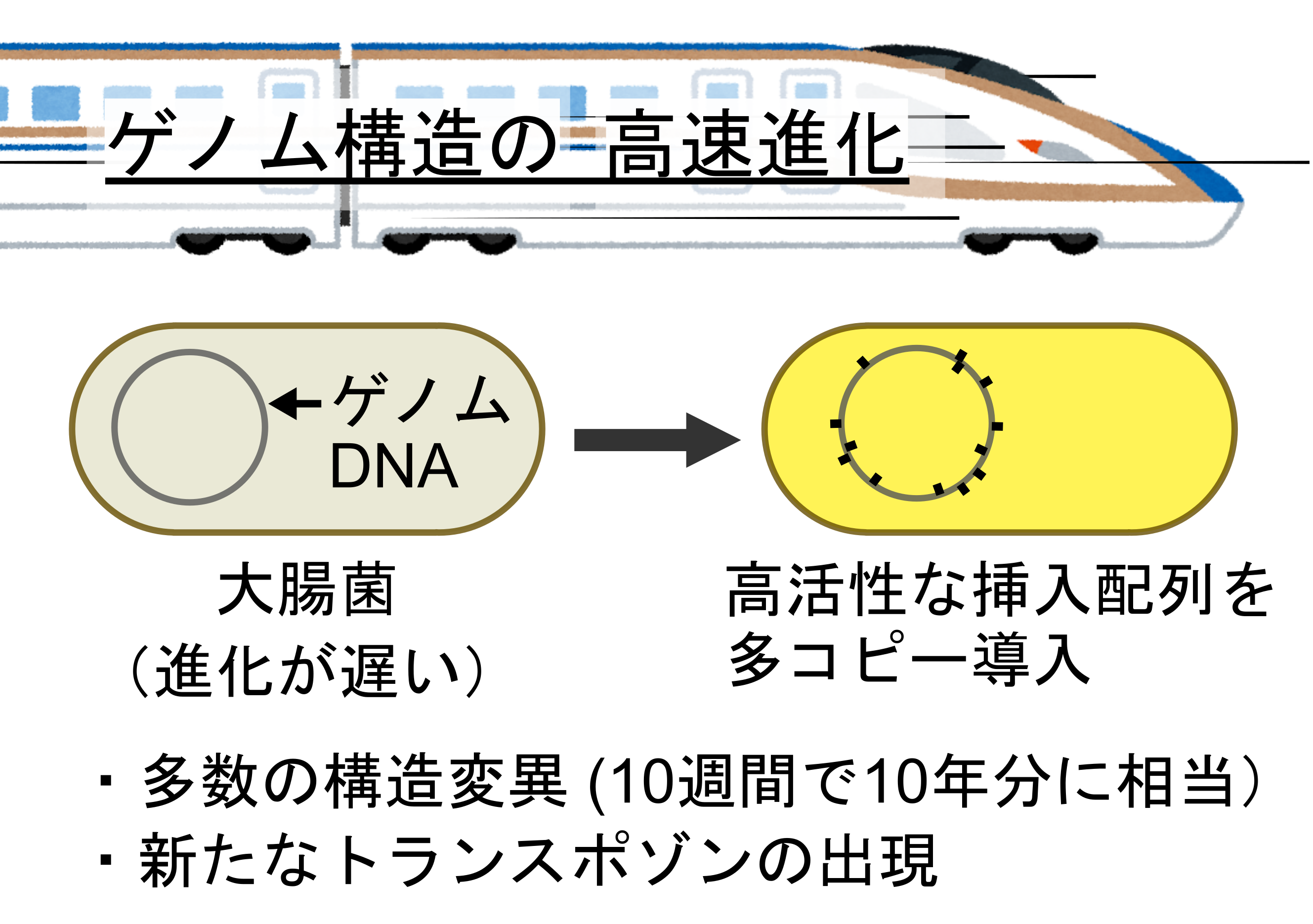
挿入配列(トランスポゾン)の導入による高速進化の概略
発表概要
東京大学大学院総合文化研究科の金井雄樹特任助教(研究当時:東京大学大学院理学系研究科大学院生)と大学院理学系研究科の津留三良特任助教と古澤力教授らによる研究グループは、細菌のゲノム構造(注1) の進化を加速する方法を開発しました。
生物の進化は点変異(DNA上の小さな変化)と構造変異(DNAの大きな変化、(注2) )によって進みます。細菌において、これまで点変異を人為的に速く起こす方法はありましたが、構造変異を速く起こす方法は限られていました。本研究では、高活性なトランスポゾン(注3) を大腸菌ゲノムに導入することで、わずか10週間で通常10年以上かかる構造変異に匹敵する大きなゲノム構造の変化を起こすことができました。さらに、進化の過程でトランスポゾンの変異体や複合トランスポゾン(図1)の出現も捉えることができました。この成果は、有用な微生物の開発や、共生細菌や病原性細菌がどのように進化するかの理解に役立つと期待されます。
発表内容
生物の遺伝情報はDNAの塩基配列として刻まれており、その進化は、DNA上の点変異と構造変異によって進みます。これまで細菌における点変異を人為的に速く起こす方法はありましたが、構造変異を速く起こす方法は限られていました。
自然界では、共生細菌や病原性細菌などの細菌が「挿入配列」と呼ばれるトランスポゾンを獲得することで、宿主や抗生物質に急速に適応することがあります。挿入配列は、自身が転移して増幅するのみならず、周りの配列の欠失・重複・逆位を引き起こします。さらに、対になって他の遺伝子を挟み、複合トランスポゾンと呼ばれる状態でまとまって転移することがあります(図1)。しかし、こうした変異の頻度が高すぎると細菌の生育に悪影響を及ぼすため、一般に挿入配列は変異が稀にしか起きないよう、活性が低くなるように進化しています。そのため、挿入配列による多数の変異が引き起こすゲノム構造の変化の軌跡を実験環境下で捉えるには、10年以上の歳月をかける必要があるのが現状でした。
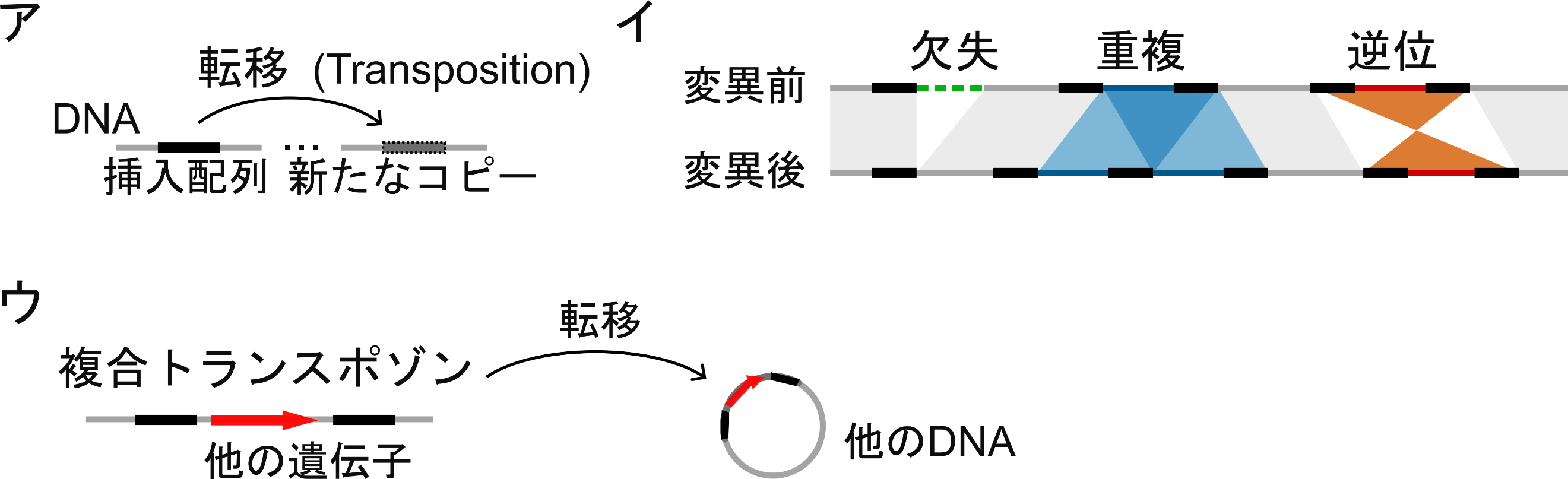
図1:挿入配列(トランスポゾン)が引き起こす構造変異
ア:挿入配列はゲノムDNA上を動く配列です。新たなコピーをゲノム上の別の位置に挿入して増えます。イ:挿入配列(黒太線部)がゲノム上にあると配列の欠失・重複・逆位といった構造変異が生じます。変異前後の対応する配列を帯で結びました。ウ:一対の挿入配列が他の遺伝子を挟み、複合トランスポゾンとして、まとめて他の位置に転移することがあります。
本研究グループは以前の研究で、挿入配列の転移を駆動するトランスポザーゼ遺伝子を改変し、さらには発現量を増やすことで、挿入配列が構造変異を起こす活性を高め、挿入配列に駆動されたオペロン(注4) 構造の進化を実験室で実証していました (Kanai et al., Nucleic Acids Research, 2022)。本研究では、同様に活性を高めた別の種類の挿入配列を大腸菌ゲノムのランダムな位置に10コピー以上導入し、44の独立した系列で10週間継代培養し、進化させました。ゲノム構造の進化過程を追跡するため、実験中の3つの時点(進化前、4週間後、10週間後)でのゲノムを長鎖DNAシーケンシング(注5) とで解読しました。そして、多数のゲノムを比較し、挿入配列が引き起こした、のべ1000を超える構造変異を自動的に検出するプログラムを開発して解析を進めました。その結果、わずか10週間で生じたゲノム構造の変化は、実験室でよく使われている大腸菌が10年以上かけて蓄積する変化に匹敵することが明らかになりました。今回開発した進化を高速化する手法により、多数の構造変異を伴ってゲノム構造が変化する過程を現実的な時間スケールで捉えられるようになったとともに(図2)、従来は断片的にしか理解されていなかった細菌ゲノムの構造変異の大きさの分布や変異の種類毎の相対的な頻度をより明確に示すことができました。
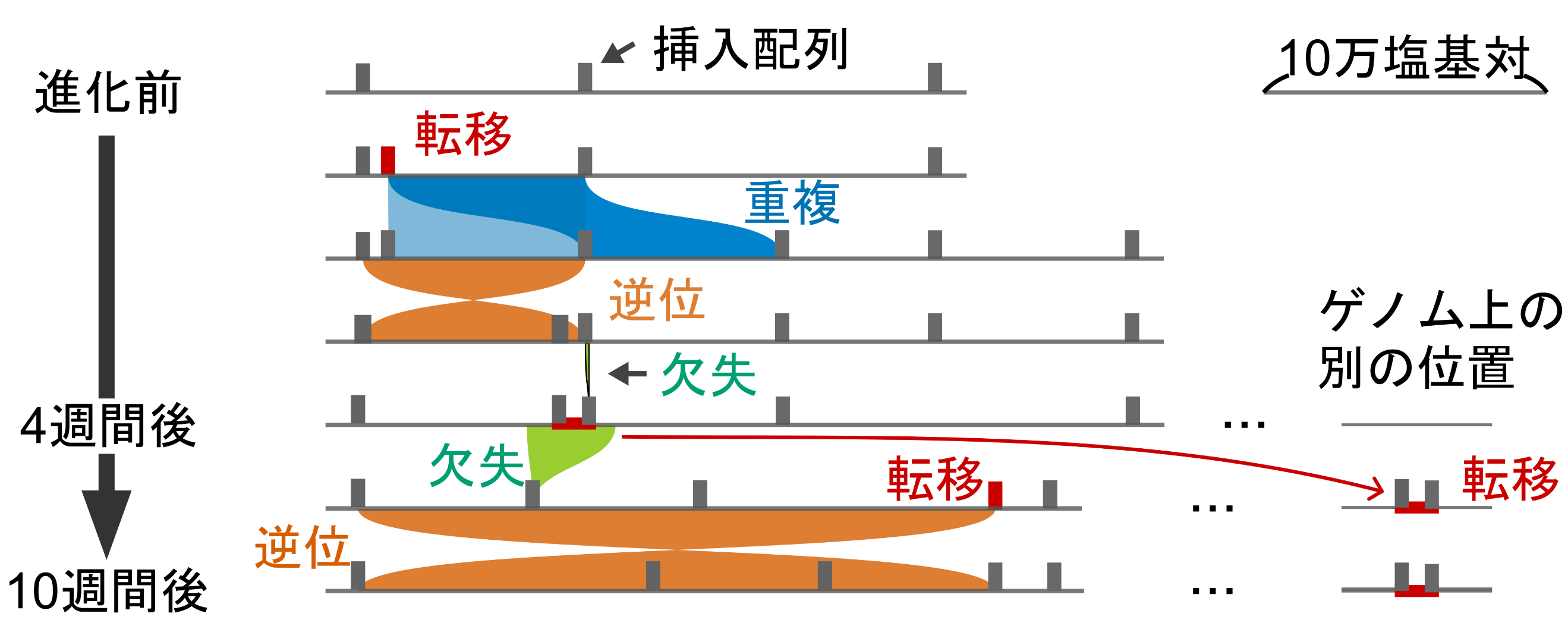
図2:挿入配列(トランスポゾン)が引き起こす構造変異
進化前、進化4週間後、および進化10週間後の大腸菌のゲノム配列を決定し、その間に起きたと推測される構造変異の順序を図示しました。図の横線がゲノム配列、ゲノム配列上の縦線が挿入配列を示します。
また、予想外なことに挿入配列同士が融合したり、挿入配列の中に挿入配列が転移し入れ子構造を形成したりすることで、新たな挿入配列の変異体が生じる過程を追うことができました(図3)。さらに驚くべきことに、多数の遺伝子が複合トランスポゾンの一部として増幅する現象を複数回観測できました。
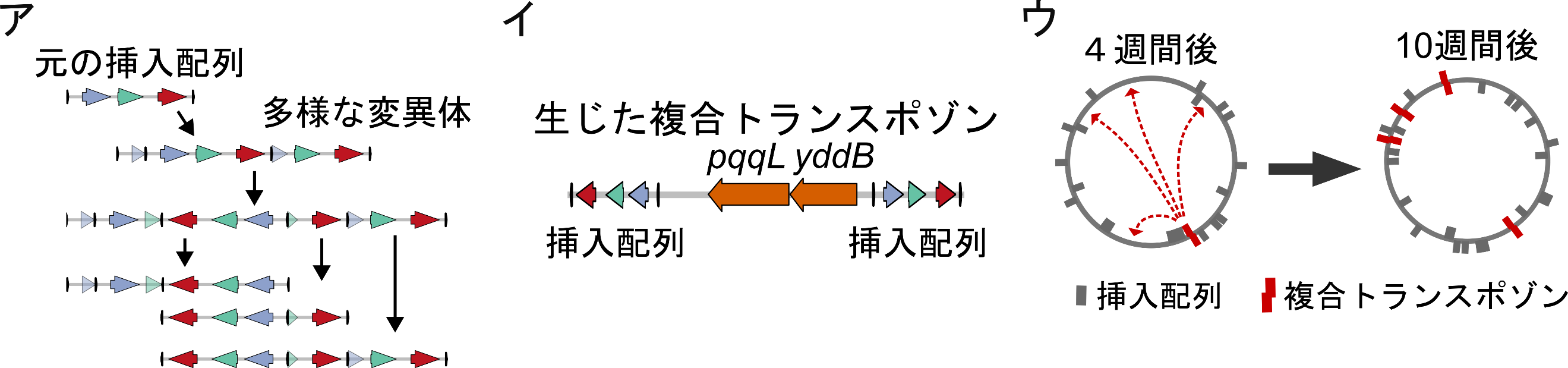 図3:挿入配列の変異体と複合トランスポゾンの出現
図3:挿入配列の変異体と複合トランスポゾンの出現
ア:初期に導入した挿入配列には3つの遺伝子(tpn、tetR、rfp)が含まれていました(左上)。進化の過程で、これらの遺伝子の配置が異なる様々な変異体が出現しました。イ:進化の過程で生じた複合トランスポゾンの例。一対の挿入配列の間に2つの遺伝子が挟まれています。ウ:進化開始4週間の時点ではこの複合トランスポゾンは1カ所にのみ存在していましたが、進化10週間までの間にゲノム上の4カ所に転移しました。
以上の6年にわたる研究の成果は、自然界では長い時間をかけて起きる細菌のゲノム構造やトランスポゾンの進化を実験室で再現し、その過程を追うことで実験科学として調べられるようにした点で重要です。本技術のさらなる発展は、産業上有用な細菌の進化工学(注6)による開発の高速化や、トランスポゾンの活性が高い共生細菌や病原性細菌の進化過程の解明に寄与することが期待されます。
関連情報
「プレスリリース オペロン構造の進化過程の実証実験に成功」(2022/01/24)
論文情報
-
雑誌名 Nucleic Acids Research 論文タイトル Laboratory evolution of the bacterial genome structure through insertion sequence activation著者 Yuki Kanai, Atsushi Shibai, Yokoi Naomi, Saburo Tsuru*, Chikara Furusawa*(*責任著者) DOI番号 https://doi.org/10.1093/nar/gkaf331
研究助成
本研究は、日本学術振興会科学研究費助成事業「特別研究員奨励費」(課題番号:21J20693、研究代表者:金井雄樹)、「基盤研究(B)」(課題番号:18H02427、研究代表者:津留三良)、「挑戦的研究(萌芽)」(課題番号:24K21985、研究代表者:津留三良)、「国際共同研究加速基金(国際先導研究)」(課題番号:22K21344、研究分担者:古澤力)、「基盤研究(B)」(課題番号:23K27164、研究代表者:古澤力)、「学術変革領域研究(A)」(課題番号:24H01798、研究代表者:古澤力)、科学技術振興機構(JST)戦略的創造研究推進事業 ERATO「深津共生進化機構プロジェクト」(課題番号:JPMJER1902,副研究総括:古澤力)、日本医療研究開発機構(AMED)の課題番号「JP223fa627001」、公益財団法人発酵研究所研究助成(課題番号:G-2025-2-046、研究代表者:古澤力)の支援により実施されました。
用語解説
注1 ゲノム構造
生物の遺伝情報は主にDNAに書かれています。そのDNAの長さやDNA分子中の遺伝子の配置のことを指します。↑
注2 構造変異
DNA上で起こる大規模な変化。配列の欠失・重複・逆位・転移などがあります。↑
注3 トランスポゾン
「動く遺伝子」の一種。ゲノム上の位置を移動するDNA配列であり、ときにコピーを作って別の場所に挿入して増えます。↑
注4 オペロン
DNAの配列情報として書かれた遺伝子は、プロモーター配列からメッセンジャーRNAへと転写され、さらにタンパク質に翻訳されて機能を発揮します。複数の遺伝子が単一のプロモーター配列からメッセンジャーRNAへと転写されるDNA配列の構造をオペロンといいます。↑
注5 長鎖DNAシーケンシング
長い断片のDNAをそのまま解読する技術。従来の短い断片を読む方法と比べて、本研究で見られるような大規模な構造変化を正確に検出できます。↑
注6 進化工学
生物の進化メカニズムを応用して、実験室での人工進化を通じて有用な微生物や酵素などを開発する方法論です。↑