
DATE2021.09.02 #Press Releases
Revealing the process of diversification of tomato leaf morphology.
Disclaimer: machine translated by DeepL which may contain errors.
Hokuto Nakayama, Assistant Professor, Department of Biological Sciences
Key points of the presentation
- Using genome information obtained with the high-speed sequencer (Note 1), we identified two causal genes for tomatoes with characteristic leaf morphology and clarified the evolutionary process of one of the genes.
- Based on these results, we have clarified the establishment process and the mechanism of leaf development for tomatoes showing characteristic leaf morphology.
- These results provide a new case study on the evolution of crop morphology at the gene level, and are also expected to lead to improvements in the morphology of crops and other plants.
Summary of Presentation
Living organisms show a wide variety of morphologies, among which crops and livestock sometimes show special morphologies that are not found in nature during the process of cultivation, domestication, and subsequent breeding. However, it is still largely unknown which genes and what kind of changes occurred during the process of domestication and breeding to produce such morphologies.
An international research team led by Assistant Professor Hokuto Nakayama of the Graduate School of Science, The University of Tokyo, and Professor Neelima Sinha of the University of California, Davis, has focused on a tomato ( Solanum lycopersicum ) variety that is grown only in certain regions and shows a characteristic leaf morphology (Figure 1). They found that changes in two different HOMEOBOX genes(Note 2) during the cultivation process caused the characteristic leaf morphology.
This research has clarified which genes and what kind of changes occurred in the process of crop cultivation that led to the diversification of morphology, and has provided knowledge that can be applied to the improvement of morphology using crops and ornamental plants in the future.
These results were published in Current Biology (online) on September 2. This research was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (JSPS), and the National Science Foundation (NSF).
Contents of Presentation
Living organisms exhibit a wide variety of forms. Among them, crops and livestock sometimes show morphologies that are not found in nature, and this is thought to be because traits that are not selected in nature are sometimes selected during the process of cultivation and subsequent breeding. Crops and livestock are therefore very interesting research subjects in terms of morphological diversity and its evolution. In fact, Charles Darwin, who brought about a new development in evolutionary studies, focused on crop and livestock diversity more than 150 years ago. However, much remains unknown about what genes and what changes have been made to crops and livestock to produce them.
To approach this question, the research group focused on tomatoes that have been grown only in certain regions, called heirloom tomatoes (Note 3), and used one of these varieties, the "Silvery Fir Tree" (SiFT), in their research. The leaves of SiFT are much more complex than those of common tomatoes (Figure 1), and its distinctive leaf shape has even led to its use as a garden and landscape plant, despite the fact that it produces edible fruit.

Figure 1: Comparison of tomato leaf morphology.
(left) M82, a strain showing common tomato leaf morphology; (right) Silvery Fir Tree, a heirloom tomato showing high complexity; seedlings were grown for 4 weeks under the same growing conditions, and the fourth leaf is used for the figure. (Bar = 2 cm).
Therefore, we first crossed SiFT, which exhibits characteristic leaf morphology, with M82, a line that exhibits common tomato leaf morphology, and the resulting F1 generation(Note 4) was self-pollinated to obtain the F2 generation. The leaves of this F2 generation showed a variety of morphologies, ranging from highly complex leaves like SiFT to those of M82. Next, we performed a genetic analysis using a high-speed sequencer, called Bulked Segregant Analysis, to identify the characteristic gene regions in the populations with high and low leaf complexity of the F2 generation. The results revealed that a portion of chromosome 2 of SiFT differs from M82 in sequence, with a single nucleotide deletion in the HOMEOBOX gene, BIPINNATA(BIP), located in that region. Using whole-mount in situ hybridization, we determined the expression site of the BIP gene and found that it is expressed at the base of the developing leaf primordium. This is the site where the lobule (Note 5) develops and is related to the complexity of the leaf. Next, we observed the leaf morphology of a mutant in which the function of the BIP gene was disrupted ( bip mutant ), and found that its complexity was higher than that of M82, and furthermore, its complexity was comparable to that of SiFT. From these results, we concluded that the BIP gene functions at the base of the leaf primordium to suppress leaf complexity, and that the deletion mutation of the BIP gene in SiFT prevents this function from being exerted, resulting in increased leaf complexity. However, when the leaves of SiFT and bip mutants were compared by image analysis using deep learning, their morphology was not the same, and SiFT had narrower leaf lobes. Other observations of the internal leaf structure also revealed that the leaf vascular bundle density was lower in SiFT than in the bip mutant. These results suggest that the BIP gene is not the only gene involved in leaf morphology in SiFT, but that another gene is also involved.
Next, to search for genes other than the BIP gene involved in the characteristic leaf morphology of SiFT, we performed RNA-seq (Note 6) using the stem apex and leaf bases of SiFT and M82, respectively, and performed a comparative analysis of the co-expressed gene network (GCN) using the data. The results revealed that SlWOX1, a homologous gene of the HOMEOBOX gene known as WUSCHEL-RELATED HOMEOBOX 1 (WOX1 ) in the model plant Arabidopsis thaliana (Arabidopsis thaliana), is the most different between the GCNs of SiFT and M82 The expression of this SlWOX1 gene was found to be the most different between SiFT and M82 GCNs. The expression level of SlWOX1 in the developing leaf primordia was found to be about half in SiFT compared to M82. The expression site of the SlWOX1 gene was also examined, and it was found to be expressed at the periphery of the developing leaf primordium. Next, examination of the leaf morphology of the mutant with disruption of SlWOX1 gene function also revealed that the lobes were narrower and the vascular bundles in the leaf were less dense than in M82. This suggests that SlWOX1, which originally has a regulatory function in controlling leaflet growth and vascular density in the width direction, is weakened by its reduced expression in SiFT, resulting in thinner leaves and lower vascular density compared to M82. In the double mutant, a cross between the bip and slwox1 mutants, leaf complexity was increased, the lobes remained narrower, and vascular density was lower. In addition, the expression level of SlWOX1 was unchanged in the bip mutant. Thus, the changes in these two genes did not interfere with each other and their phenotypes were additive. Based on the above, we conclude that in SiFT, the loss of function of the BIP gene mutation and the reduced expression of the SlWOX1 gene resulted in a unique leaf morphology with increased leaf complexity, narrower lobes, and lower vascular density (Figure 2).
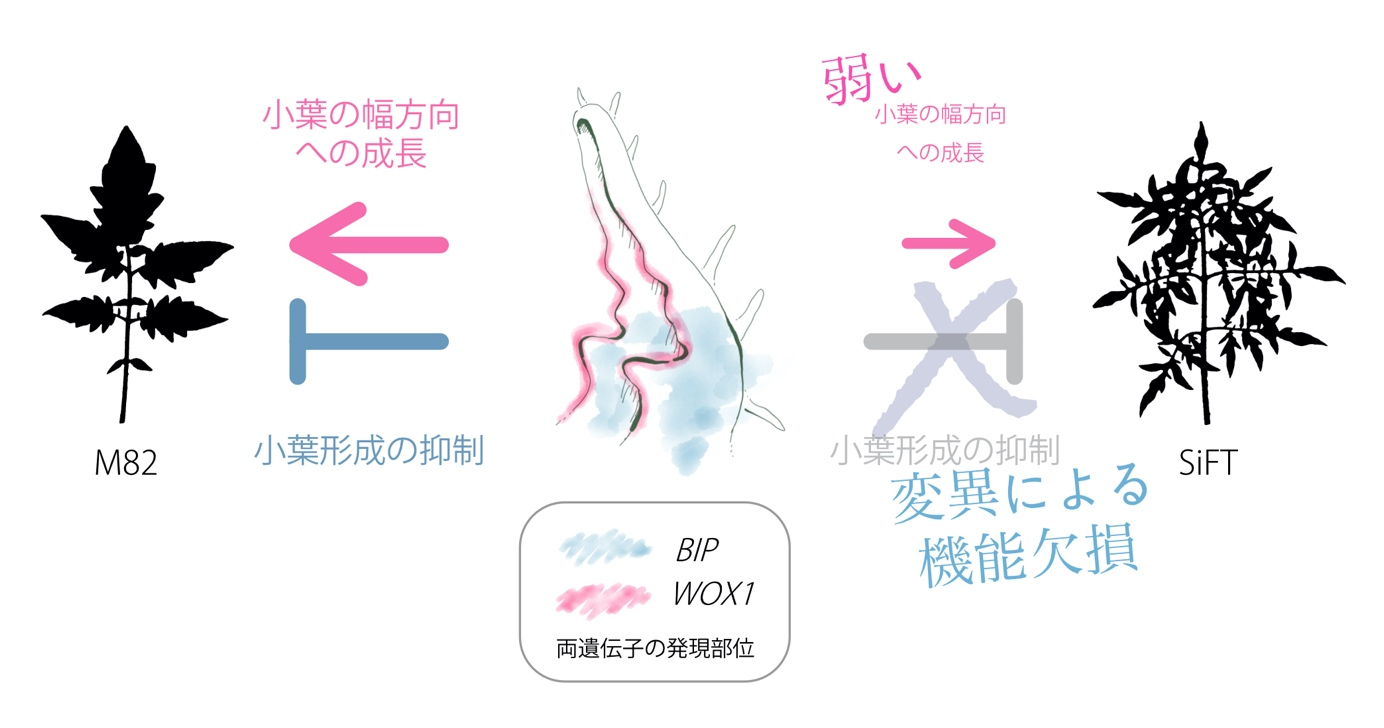
Figure 2: Mechanism of "high complexity" and "narrow" leaves in Silvery Fir Tree.
The leaf morphology found in tomatoes in general has a shape similar to that of M82, which involves a number of genes including BIP andSlWOX1. In the Silvery Fir Tree, on the other hand, the BIP gene, which suppresses the formation of lobes, has been mutated and is no longer able to suppress the formation of lobes. In addition, the function of the SlWOX1 gene, which makes each lobule grow in the width direction, is also weakened, resulting in "highly complex" and "thin" leaves.
Finally, the group also looked at how the bip mutation found in SiFT arose during evolution. To do this, they sequenced the genomes of several new heirloom tomatoes in addition to the various tomato genome data already in the database, and used these genome data to infer the evolutionary process. We found no bip mutations in the genomes of any of the tomato lines related to SiFT, including those closely related to SiFT and those suggested to be interbred with the ancestral lineage of SiFT. In fact, the leaf morphology of those tomatoes, like M82, is known to be of low complexity, and wild tomato species prior to cultivation are also known to be of low complexity. These facts suggest that the bip mutation in SiFT was newly acquired during the cultivation process and was not introduced by hybridization from other tomatoes.
These analyses indicate that the characteristic leaf phenotype of SiFT, a heirloom tomato with high complexity, narrow lobes, and low vascular density, is caused by changes in two HOMEOBOX genes, theBIP gene and theSlWOX1 gene. The BIP gene and the SlWOX1 gene were also found to be involved in the phenotype. The mutation in the bip gene was not introduced from other tomatoes, but was newly acquired during the cultivation process (Fig. 3).
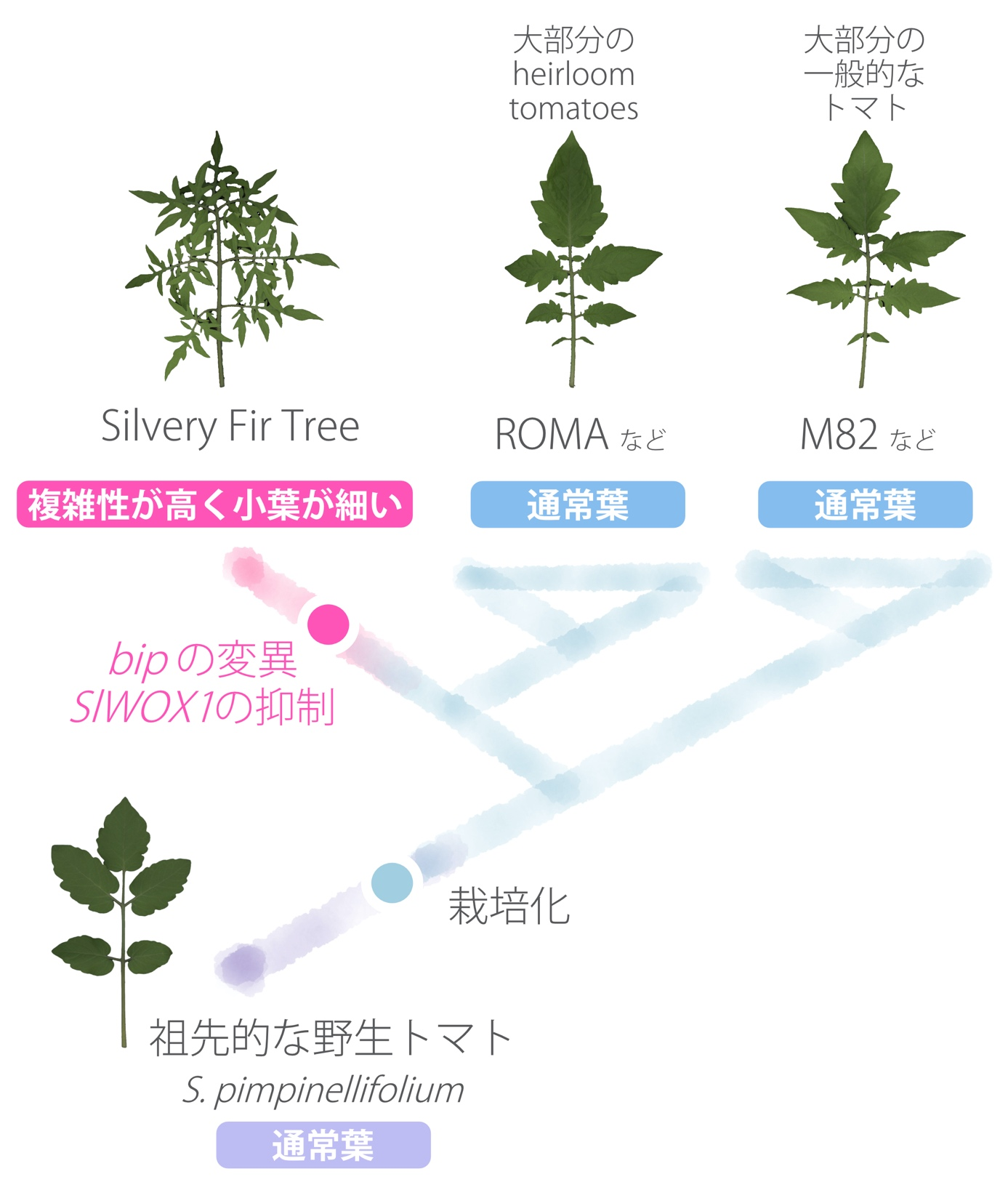
Figure 3: Silvery Fir Tree formation process in the history of tomato cultivation and breeding.
The majority of tomatoes have a leaf morphology similar to that of M82. The wild tomato, prior to cultivation, also has a leaf morphology similar to that of M82 ( Solanum pimpinellifolium is known to be the ancestor of the current tomato species). The present study indicates that the high complexity of the Silvery Fir Tree, plus the narrow width of the lobes, a characteristic leaf phenotype not seen in other tomatoes, was caused by changes in two HOMEOBOX genes, theBIP gene and theSlWOX1 gene.
Leaves are found in many plants and their morphology is very diverse. Sometimes, as in this case, special morphologies are selected during the cultivation process and are pleasing to our eyes. In addition, recent studies have shown that leaf morphology and vascular bundle density are related to the sugar content of the fruit, and it may be possible to improve the taste and nutrients of the fruit by modifying these factors. Therefore, the results of this research provide important knowledge not only from the evolutionary perspective of which genes and what kind of changes occurred in the process of crop cultivation that led to the diversification of forms, but also for the improvement of crop varieties.
Journal
-
Journal name Current Biology Title of paper Leaf form diversification in an ornamental heirloom tomato results from alterations in two different HOMEOBOX genes Author(s) Hokuto Nakayama, Steven D. Rowland, Zizhang Cheng, Kristina Zumstein, Julie Kang, Yohei Kondo, Neelima R. Sinha DOI Number 10.1016/j.cub.2021.08.023
Publication URL
Terminology
Note 1 High-speed sequencer
A device capable of decoding several hundred million to several billion bases of DNA at a time. This has made it possible to perform genome analysis and comprehensive gene expression analysis more easily than in the past. Vigorous improvements are still being made, and the total number of bases that can be decoded at one time and the length per actual sequence obtained, called reads, are increasing. ↑up
Note 2 HOMEOBOX gene
A gene whose sequence (homeobox) was conserved in the gene responsible for the homeotic mutation in Drosophila. It is found in plants, animals, and fungi and is known to be involved in the regulation of developmental processes. ↑up
Note 3 heirloom tomato
Tomatoes that have been cultivated and passed down for generations only in certain regions or, in some cases, within families. The image is similar to that of Japan's kyoyasai (Kyoto vegetables). Unlike the tomatoes that are mainly distributed today, heirloom tomatoes are very diverse in taste, color, and shape, and are favored for gardening and cooking. ↑up
Note 4 F1 generation
The first generation of a hybrid. The offspring of the first generation resulting from the crossbreeding of different parents. In the case of the second generation of hybrids, it becomes F2. ↑up
Note 5 Small leaf
A leaf like a tomato, in which one leaf is composed of multiple units (like small leaves), is called a compound leaf, and each of these units is called a lobule. The more lobes there are, the more complex the leaf. A leaf that is not classified as a compound leaf (i.e., one that is not clearly divided at the leaf margin) is called a monolobe. ↑up
Note 6 RNA-seq
RNA-seq is a method that uses a high-speed sequencer to decode transcripts in a tissue or a cell, and enables comprehensive quantification of expression levels. By statistically processing the data, genes with similar expression patterns can be extracted and their relationships can be expressed in the form of a network (called a co-expressed gene network). ↑up