
DATE2021.04.08 #Press Releases
Chloroplasts of plants tolerate low light and occasional intense light
Disclaimer: machine translated by DeepL which may contain errors.
Ichiro Terashima (Professor, Department of Biological Sciences)
Mitsutoshi Matsuo (at the time of research: 2nd year Master's student, Department of Biological Sciences)
Yoshihiro Suzuki (Associate Professor, Department of Biological Sciences, Faculty of Science, Kanagawa University)
Wataru Yamori, Associate Professor, Graduate School of Agricultural and Life Sciences
Masaru Kono, Project Assistant Professor, Department of Biological Sciences
Key points of the presentation
- Chloroplasts of the shade-tolerant plant, the mulberry, which tolerates dim light, are not easily damaged by occasional exposure to intense light, and the chloroplasts of plants grown in low light are more tolerant than those of plants grown in high light.
- Chloroplasts of plants grown in high light are more tolerant of constant high light, but when light intensity fluctuates, as in a forest where the sun shines through the trees, chloroplasts of plants grown in low light are more tolerant.
- Injury to photosynthetic organs (photoinhibition) caused by intense light is one of the major factors that reduce plant productivity. The findings of this study will provide a basis for the creation of plants tolerant to strong light injury under natural light conditions.
Summary of Presentation
Photosynthesis is a reaction that uses light energy to synthesize organic matter and is carried out in chloroplasts contained in leaf cells. Light is essential for photosynthesis, but too much intense light can damage chloroplasts. Of the photosystems I and II (Note 1 ) that convert light energy into chemical energy, it is well known that photosystem II is mainly damaged by continuous exposure to intense light, and that plants grown under intense light are more resistant to this damage.
Professor Ichiro Terashima, Graduate Student Mitsutoshi Matsuo and Project Associate Professor Masaru Kono at the Graduate School of Science, The University of Tokyo, Associate Professor Wataru Yamori at the Graduate School of Agriculture, The University of Tokyo, and Associate Professor Yoshihiro Suzuki at the Department of Biological Sciences, Kanagawa University, and their colleagues exposed several types of plants to variable light and compared the damage. They found that the photosystem I of the Japanese knotweed, which grows even under low light conditions, is not easily damaged, and that the lower the light intensity during cultivation, the more resistant to "variable light" the photosystem I is ( Fig. 1). (Figure 1)

Figure 1: A wolverine potato in the sunlight shining through a tree. Photographed in a field on the campus of The University of Tokyo.
To search for the cause of this phenomenon, we analyzed chlorophyll fluorescence in leaves, the redox state of the photosystem I reaction center, various spectra of isolated chloroplasts, and the composition of chlorophyll protein complexes. These results indicate that photosystem I is well excited in low-light-grown quail and that the amount of electrons flowing from the photosystem II side into the photosystem I reaction center is suppressed under high light. The reaction center of photosystem I was in an oxidized state with a loss of electrons because it was well excited even though the amount of electrons flowing into the reaction center was small. The charge recombination (Note 3 ) between the oxidized reaction center and the reduced electron carrier is thought to scavenge excess energy.
Photoinhibition is a bottleneck in plant photosynthetic production, and attempts are being made to create photoinhibition-resistant plants. The findings obtained in this study are key to the creation of photoinhibition tolerant plants under natural conditions.
Content of Presentation
Photosynthesis is a reaction that uses light energy to synthesize organic matter. Photosynthetic products of plants and algae nourish not only plants and algae but also most living organisms on the earth. Photosynthesis in plants takes place in chloroplasts contained in leaf cells. When light excites chlorophyll in the protein complexes of chloroplasts, the excited state is transferred from one chlorophyll to another and finally to the reaction center chlorophyll in the photosystem. When the reaction center chlorophyll is excited, electrons leave the reaction center chlorophyll and initiate a series of redox reactions. Thus, light is an essential energy source for driving photosynthetic reactions. On the other hand, too much light energy is used to generate reactive oxygen species such as singlet oxygen and superoxide (Note 4), which damage chloroplasts. There are two types of photosystems: photosystem II, which oxidizes and decomposes water with strong oxidative power, and photosystem I, which produces stable reducing agents. It has also been shown that plants grown under high light intensity are more tolerant to damage caused by high light intensity.
Project Assistant Professor Masaru Kohno and his colleagues at the Graduate School of Science, The University of Tokyo, have been pursuing the cause of the damage to photosystem I, but not photosystem II, under "variable light," which is often observed in outdoor natural light environments, where plants are occasionally exposed to high light in addition to low light. We also found that in the presence of far-red light, which drives only photosystem I, this inhibition is almost non-existent. We compared the degree of damage to photosystem I by variable light in several species, and found that photosystem I in the shade-tolerant quail, which can grow in low light, was tolerant of variable light even in the absence of far-red light. Moreover, it was found that the more tolerant the quatrain I was grown under low light, the more tolerant it was. (Figure 2)
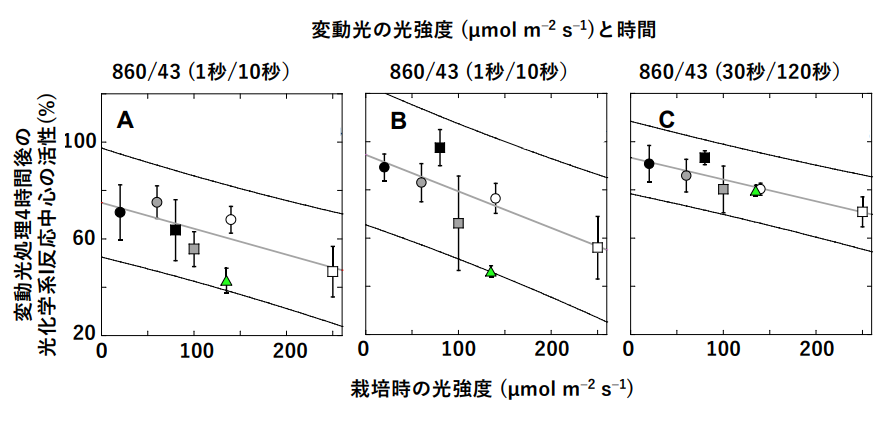
Figure 2: Effect of variable light treatments and light intensity during cultivation on the activity of photosystem I.
Three variable light treatments alternating between high and low light were applied for 4 h. 100% light was much stronger than cultivated light with a photosynthetically effective photon flux density (the amount of photons falling at wavelengths of 400~700 nm) of 860 µmol m-2 s-1. This is about 40% of the photosynthetically effective photon flux density during the daytime in midsummer. Plants were grown under 12 hours of continuous light. The reaction center (P700) of photosystem I is oxidized when exposed to light. The vertical axis of the figure shows the ratio of the amount of P700 oxidized by light after the variable light treatment to that before the variable light treatment. Data indicated by □ and ○ in each panel are means and standard deviations for leaves of five individuals grown at each light intensity. The green △ indicates data for the model plant Arabidopsis thaliana. Regression lines and 95% confidence intervals for the data on the quail potato are also shown.
To investigate the cause of this phenomenon, chlorophyll fluorescence in leaves and the redox state of the photosystem I reaction center were examined. We also measured various spectra of isolated chloroplasts and quantified the photosystem I reaction center and cytochrome f(Note 5), an electron transfer component. We also analyzed the composition of chlorophyll protein complexes. From these results, it became clear that the following two factors were responsible for the resistance of photosystem I of low-light-grown plants to photoinhibition by fluctuating light: 1) the photosystem I is more resistant to photoinhibition than the photosystem I of low-light-grown plants, and 2) the photosystem I is more resistant to photoinhibition than the photosystem I of low-light-grown plants. (1) the amount of electrons flowing from photosystem II to photosystem I via cytochrome f during high light in variable light is low because the amount of photosystem I is high compared to that of photosystem II and cytochrome f. (2) during high light, the light energy absorbed by the leaves is distributed more to photosystem I. (3) during low light, the light energy absorbed by the leaves is distributed more to photosystem I. (4) the amount of electrons flowing from photosystem I to photosystem II is low compared to that to photosystem II. (2) In high light conditions, more light energy absorbed by the leaf is distributed to photosystem I. When the amount of electrons flowing into photosystem I is small, but a large amount of excitation energy is distributed to photosystem I, the reaction center in photosystem I loses electrons and enters an oxidized state. Charge recombination between this and the electron transfer component in the reduced state appears to successfully scavenge the excess energy. (Figure 3)
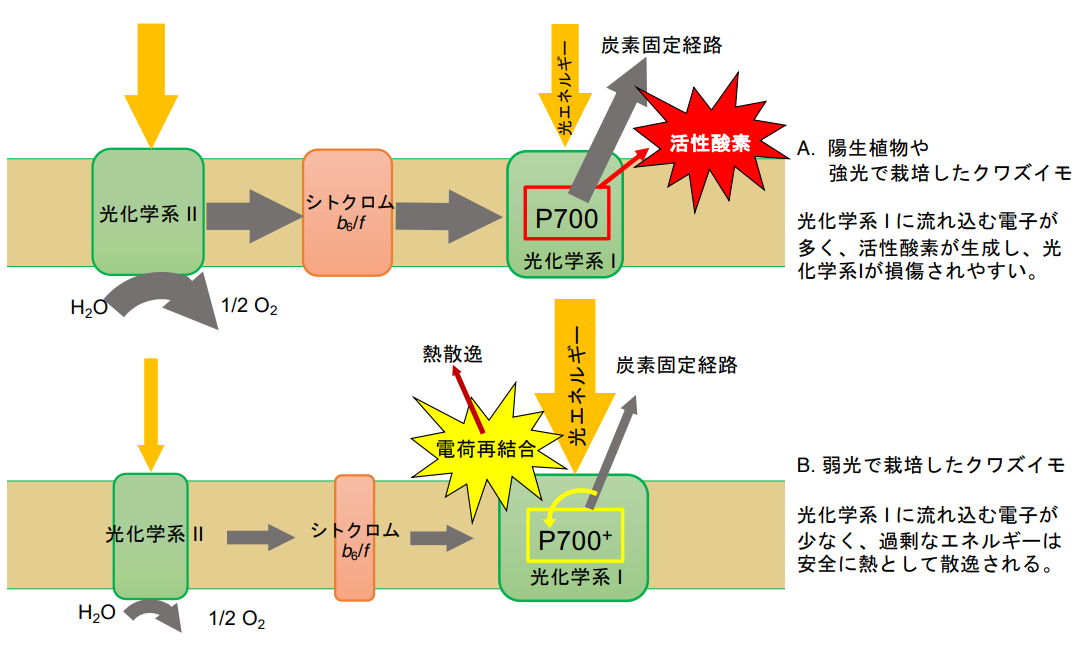
Figure 3: Photosynthetic membranes in variable light conditions, from low light to high light.
A. In the case of a yang-grown plant or arabidopsis grown under high light: A large amount of electrons flow into photosystem I from photosystem II and cytochrome complexes containing cytochrome f . The electrons flow into photosystem I from photosystem II and cytochrome f . The electrons flow into photosystem I from cytochrome complexes containing cytochrome f . If the amount of electrons required to drive the CO2 fixation pathway is less than the amount of electrons flowing into photosystem I, electrons flow into O2 to form superoxide, which causes photosystem I is damaged.
B. In the case of quails grown under low light: The electron flux flowing from photosystem II to photosystem I via cytochrome f is small, and photosystem I is well excited. The reaction center of photosystem I is in the oxidized state ( P700+), and charge recombination occurs with the electron transfer component in the reduced state. During charge recombination, the excess energy in photosystem I is safely dissipated as heat, so photodamage is unlikely to occur.
Photoinhibition suppresses plant photosynthetic production, and the creation of photoinhibition tolerant plants is thought to lead to increased crop production. The results of this study strongly demonstrate the need for research on photoinhibition of photosystem I and its tolerance, rather than just focusing on photosystem II, which has been the focus of previous studies.
Journals
-
Journal name Photosynthesis Research Title of paper Photosystem I in low light-grown leaves of Alocasia odora, a shade-tolerant plant, is resistant to fluctuating light-induced photoinhibition Authors Ichiro Terashima*, Mitsutoshi Matsuo, Yoshihiro Suzuki, Wataru Yamori, Masaru Kono*, and Masaru Kono DOI Number https://doi.org/10.1007/s11120-021-00832-4 URL https://link.springer.com/article/10.1007/s11120-021-00832-4
Terminology
1 Photosystem I and II
Chlorophyll is excited by light energy, and the excited state is transferred from one chlorophyll to another between neighboring chlorophylls, eventually arriving at a chlorophyll dimer called the reaction center. When the reaction center chlorophyll is excited, electrons leave the reaction center and reduce a series of electron transfer components. The reaction center, which has lost an electron, exhibits strong oxidizing power. Photosystem II uses its oxidizing power to oxidize water and steal electrons. Electrons from photosystem I reduce NADP+ by two electrons with the electrons gained from water, producing a stable reducing agent (NADPH) used in biochemical reactions ( NADP++H+ +2 e- →NADPH). ↑ (NADPH)
Note 2 shade tolerance
Plants that prefer brightly lit areas are called xerophytes, while those that prefer dimly lit areas are called shade-tolerant. Although it is difficult to call the kwazuiimo a shade plant because it flowers in bright locations such as forest edges, it is a shade-tolerant plant that can tolerate fairly dark locations and is used as an indoor houseplant. It is a member of the taro family, the same as Monstera and Pothos. ↑up
Note 3 Charge recombination
The reaction center of photosystem I in the reduced state is called P700 because it shows absorption at 700 nm. When charge recombination occurs between the oxidized P700+ and the electron transfer component ( A-) in the reduced state near P700, P700+ +A- → P700 + A. The energy generated at this time is dissipated as heat. If the energy produced at this time is dissipated as heat, then idling is safe. ↑up
Note 4: Reactive oxygen species
Superoxide ( O2-), a one-electron reduction of oxygen (O2 ), and singlet oxygen (singlet oxygen, 1O2 ), in which excited chlorophyll enters a triplet state that excites O2. Both have strong oxidizing power and destroy biological substances. ↑up
Note 5 Cytochrome f
Electron transfer electron between photosystem II and I. Steady-state photosynthetic rate is modulated to the same degree by several photosynthetic components. One of these is cytochrome f. The maximum photosynthetic rate (generally expressed per leaf area) obtained at steady-state high light intensity is strongly correlated with the amount of cytochrome f ↑up