
DATE2021.07.29 #Press Releases
水陸両生の水草ミズハコベが姿を変える仕組みを解明
古賀 皓之(生物科学専攻 助教)
小嶋 美紀子(理化学研究所 専門技術員)
竹林 裕美子(理化学研究所 テクニカルスタッフ)
榊原 均(名古屋大学 教授/理化学研究所 客員主管研究員)
塚谷 裕一(生物科学専攻 教授)
発表のポイント
- オオバコ科の水陸両生植物(注1) ミズハコベが、水中と陸上とで葉の形を変える現象「異形葉性」の分子基盤を明らかにした。
- ミズハコベの異形葉性には複数の植物ホルモンが関わっていることを明らかにし、さらにその下流で葉の形態の制御に重要な役割を果たす遺伝子群を絞り込んだ。
- 異形葉性は複数の被子植物の系統で独立に進化しており、ミズハコベの系統においても、独自の仕組みで異形葉性が進化してきたことが示唆された。植物の水辺環境への適応の仕組みを知る手がかりとなる。
発表概要
水辺に生育する植物である水草の仲間には、水中でも陸上でも育つことのできる水陸両生の種が知られています。水陸両生種ではしばしば、同一個体でも、陸上で成長している時と水中で成長している時とで、劇的に異なる形態の葉を作ることが知られています。この能力は異形葉性と呼ばれ、古くから研究者の興味を引いてきました。しかし、どうして全く異なる葉の形を作ることができるのか、その仕組みについては詳しく知られていませんでした。東京大学大学院理学系研究科生物科学専攻の古賀 皓之助教、塚谷 裕一教授を中心とする本研究グループは、顕著な異形葉性を示すオオバコ科の水草ミズハコベを用いて、異形葉性の制御機構について研究を進めました。
ミズハコベは陸上では卵形の陸上葉を、水中ではリボン状の水中葉をつけます。この水中葉は、葉を構成する各細胞が細長く成長することで作られます。本研究グループは、水中葉の細胞成長には植物ホルモンのエチレン(注2) やジベレリン(GA)(注3) の作用が必要であることを示しました。また一方で、別の植物ホルモンであるアブシシン酸(ABA)(注4) の内生量の減少も必要であることがわかりました。しかし、これらのホルモンの変化だけでは、陸上で成長している植物に水中葉を作らせることはできないこともわかりました。水中葉の形成には、水中にいることで引き起こされる、なんらか別の因子も必要なことが示唆されます。
さらに研究グループは、複数の栽培条件での遺伝子発現パターンを網羅的に比較した上、加えて異形葉性をもたない近縁種とも遺伝子発現パターンを比較することで、異形葉性に深く関わる遺伝子群を絞り込みました。これらの遺伝子のうちいくつかは、モデル植物シロイヌナズナで細胞伸長に関与することが知られているものでした。こうした遺伝子群が複合的に働くことにより、劇的な葉の発生様式の変化を引き起こしていると推定されます。
この成果は、米科学誌『The Plant Cell』で公開されました。
発表内容
水中でも陸上でも育つことのできる水陸両生植物は、陸上に生えているときには陸上での生存に適した形の葉を、水中に生えているときには水中に適した形の葉を作ることができます。このように、同一の個体が異なる環境で全く異なる形の葉を作る能力を、異形葉性と呼びます。水陸両生植物の異形葉性は非常に劇的な変化を示すので、古くから研究者の興味を引いてきました。しかしその分子的な仕組み、例えばどのような遺伝子の作用によってこのような葉の形の変化が引き起こされるのか、といった点については最近まで研究が進んでいませんでした。近年いくつかの研究グループが、別々の水草を使って異形葉性の分子機構の解明を進めていますが、詳しい仕組みの理解には至っていません。また、異形葉性の進化は植物のさまざまな系統で独立に起こったことが知られており、ある系統の水草でわかった仕組みが、他の系統の水草にもあてはまるのかどうか、といったこともよく分かっていませんでした。そこで今回、研究グループは、オオバコ科の水草ミズハコベを使って、本種の異形葉性の分子機構を調べました。
ミズハコベは、日本でも湧水河川などでよくみられる水草です。小型で扱いやすく、研究室内で容易に育ち、また非常に劇的な異形葉性を示すため、その分子基盤を調べるのに好適な材料として、筆頭著者らによって研究基盤が整えられてきました。ミズハコベは陸上では卵形の葉を作るのに対し、水中では細長いリボン状の葉を作ります(図1)。

図1:ミズハコベの異形葉性
ミズハコベ(学名: Callitriche palustris)を陸上環境で育てたときの姿(左)と水中環境で育てたときの姿 (右)。どちらも同一の植物から挿し木したクローンである。
このリボン状の葉は、葉を構成する各細胞が非常に細長く成長することで作られます。このとき、植物内では植物ホルモンのエチレンとジベレリン(GA)が働いていることがわかりました。これらのホルモンの作用を阻害する薬品を加えて育てると、水中であっても陸上葉のような葉を作ることが明らかになったからです(図2)。また、別の植物ホルモンのアブシシン酸(ABA)を加えても水中葉は形成されず(図2)、実際に水中ではABAの内生量が減少していたため、水中葉を作るためにはABAの合成を抑える必要があることがわかりました。
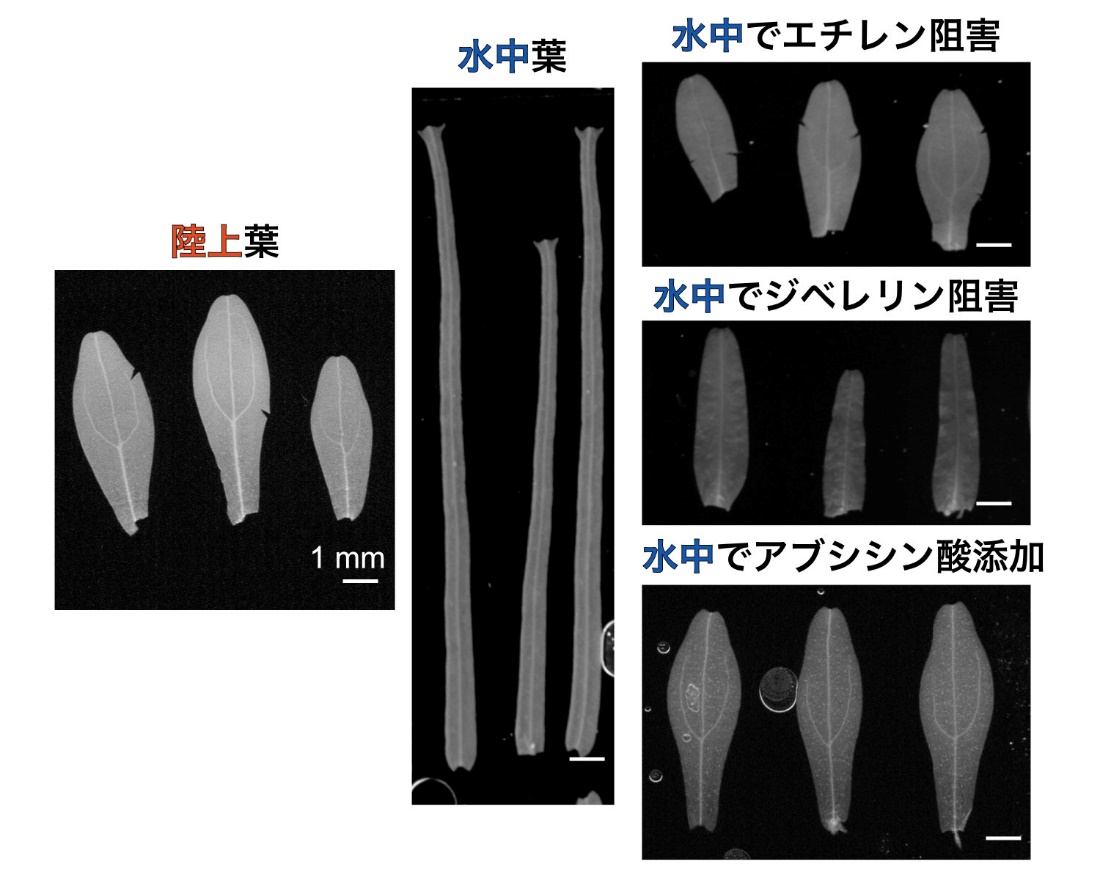
図2:植物ホルモンやその阻害剤の作用
ミズハコベは陸上では卵形の陸上葉を、水中では細長い水中葉を作る。しかしエチレンシグナルの阻害剤やジベレリン生合成の阻害剤、またはアブシシン酸(ABA)を加えて育てると、水中でも陸上葉に近い形の葉を作る。
しかし逆に、陸上でエチレンとジベレリンを与えても、葉の形の変化はごくわずかであり、これらのホルモンの作用だけでは水中葉を形成できないことが示唆されました。これまで先行研究で調べられてきた水草では、エチレン添加によって陸上環境でも劇的な形態の変化が誘導できるとされてきたことと、対象的な結果です。ミズハコベの水中葉形成を引き起こすには、植物ホルモンの変化の他に水中環境と関連した別の因子の作用も必要だと考えられます(図3)。
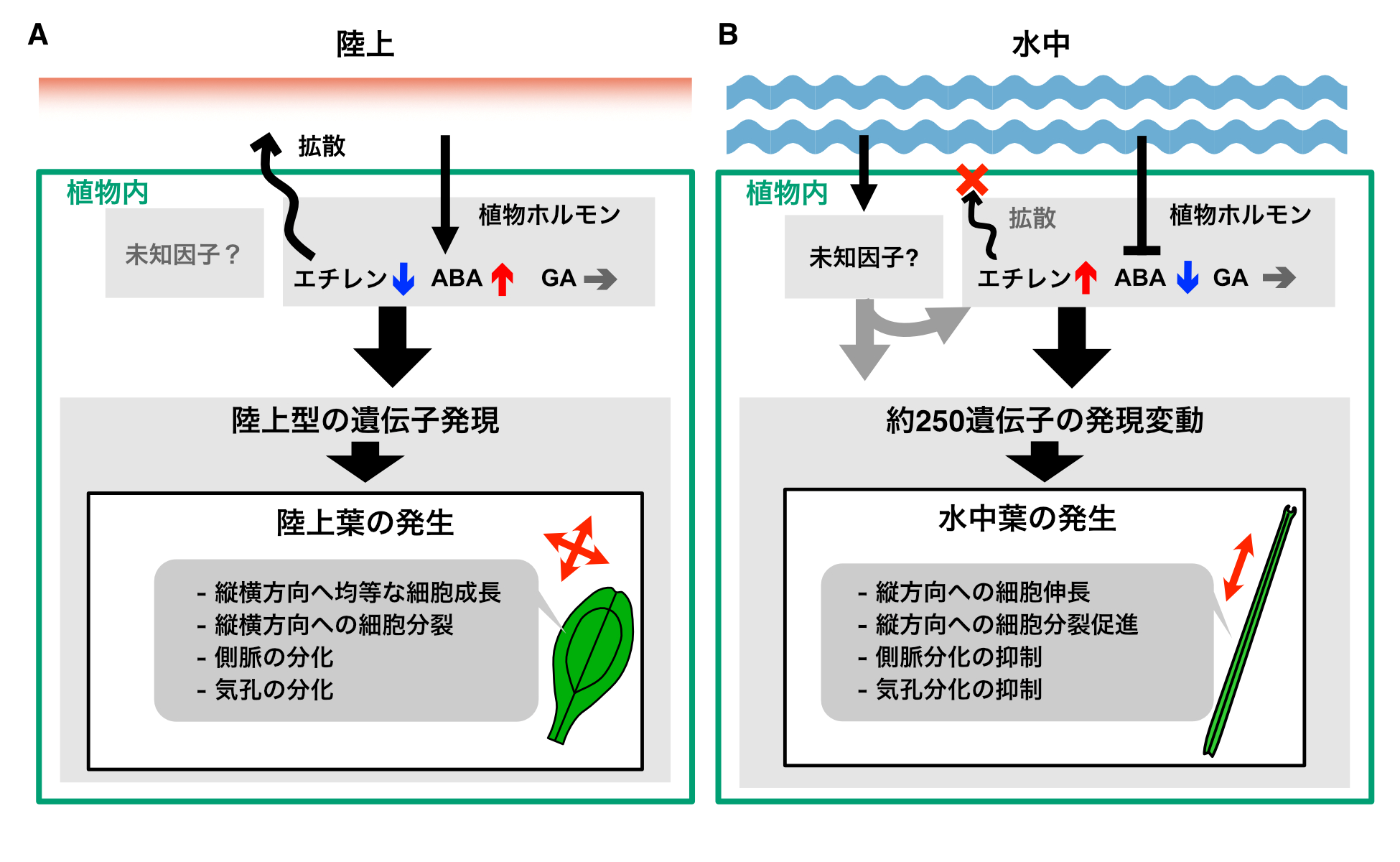
図3:ミズハコベにおける異形葉性制御の概要
(A)陸上では拡散によるエチレンシグナルの減衰や、アブシシン酸(ABA)の増加、そしてその状況下でおこる遺伝子発現によって陸上葉が発生する。(B)水中ではエチレンの蓄積やABAの減少、さらに何らかの因子の影響によって遺伝子発現に変化が起こり、水中葉が発生すると考えられる。ジベレリン(GA)の内生量は変化しないが、その作用は水中葉の形成に必要である。
研究グループはさらに、トランスクリプトーム解析(注5)を用いて、陸上と水中、および植物ホルモン阻害下で、発生中の葉における網羅的な遺伝子発現パターンを明らかにしました。また、近縁種ながら陸生で、異形葉性を持たないアメリカアワゴケでも遺伝子発現パターンを明らかにし、それらのデータも比較することで、ミズハコベでの異形葉性の制御に深く関わる遺伝子群を絞り込みました。250個ほどに絞られた遺伝子の変化には、水中葉では抑制される気孔(注6)形成に関わる遺伝子の発現減少や、モデル植物のシロイヌナズナ等で、細胞伸長を促進することが知られている遺伝子の発現上昇などが検出されました。しかし、これらすべての遺伝子は、ミズハコベにおいてどのような働きをしているのかが全く分かっていません。今後ミズハコベを用いて、ゲノム編集等の技術を用いた遺伝子機能解析等を行なうことで、こうした遺伝子が、異形葉性にどのように関わるのかを明らかにしていく必要があります。
現在地上に繁栄している植物は、かつて水中から陸上に進出しました。そして、水草はそこから再度水辺や水中へもどっていった植物です。水草の異形葉性は数多くの系統で独立に進化してきましたが、多くの場合水中葉の形態は、陸上葉とくらべて顕著に細長いという点で共通しており、収斂進化(注7)の例として知られています。今回の研究結果からわかったミズハコベの異形葉性の仕組みを、これまでに研究されてきた水草と比べてみると、関わっている植物ホルモンこそ共通していますが、その作用の仕方や、下流で働く遺伝子群は大きく異なることが示唆されました。これは、水陸両生植物は系統によって独自のやり方で、水中の環境に適した細長い葉を作る仕組みを実現したことを反映したものと思われます。今後、こうした仕組みをさらに詳しく調べることで、植物の世界では決して珍しくはない、水辺環境への再進出という進化イベントの、全容の解明につながると考えられます。また、植物の水辺環境への適応の仕組みを知ることで、洪水等に強い作物を作ることにも繋がるでしょう。
発表雑誌
-
雑誌名 The Plant Cell 論文タイトル Identification of the unique molecular framework of heterophylly in the amphibious plant Callitriche palustris L. 著者 Hiroyuki Koga*, Mikiko Kojima, Yumiko Takebayashi, Hitoshi Sakakibara, Hirokazu Tsukaya DOI番号 10.1093/plcell/koab192
論文URL
用語解説
注1 水陸両生植物
水辺に生育する植物(水草)の生活型のひとつ。完全水生の水草は水中では成長できるが、陸上では乾燥して死んでしまう。また、通常の陸生植物は、水中では成長が著しく阻害され、生存できない。しかし水陸両生の水草は、水中でも陸上でも成長することができるため、水位が頻繁に変動するような環境で有利に生存できると考えられる。 ↑
注2 エチレン
植物ホルモンの一種で、常温で気体。種子の発芽促進、茎の伸長抑制、果実の成熟促進、花や葉の老化・脱落促進などのはたらきがある。気体であるため、水中では植物体外への放出が著しく阻害される。 ↑
注3 ジベレリン(GA)
ある種の植物ホルモンの総称で、一般に生長軸の方向への 細胞伸長促進、種子の発芽促進や休眠打破の促進、 老化の抑制等に関わっている。↑
注4 アブシシン酸(ABA)
植物ホルモンの一種。休眠や生長抑制、気孔の閉鎖などを誘導する。 また乾燥などのストレスに対応して合成されることから「ストレスホルモン」とも呼ばれる。 ↑
注5 トランスクリプトーム解析
超並列シーケンサーなどを用いて、遺伝子の転写産物(トランスクリプト)を網羅的に検出する解析手法。近年の技術の発達により、遺伝子情報の乏しい生物でも、ほぼすべての遺伝子の配列情報と発現量が容易に解析できるようになった。↑
注6 気孔
植物のガス交換のための器官で、葉や茎に存在する孔。水中ではガス交換ができず、また病原菌等の侵入経路にもなりうるためか、多くの水陸両生植物は水中では気孔を作らない。↑
注7 収斂進化
共通の祖先がもっていた特徴ではないのにも関わらず、系統的に離れたグループ間で形態的または機能的によく似た特徴が進化すること。鳥類とコウモリの翼、脊椎動物とタコやイカのカメラ眼、などが収斂進化の例として挙げられる。↑